Diversification and genetic regulation of cranial neural crest cell fates across the lifetime of a vertebrate
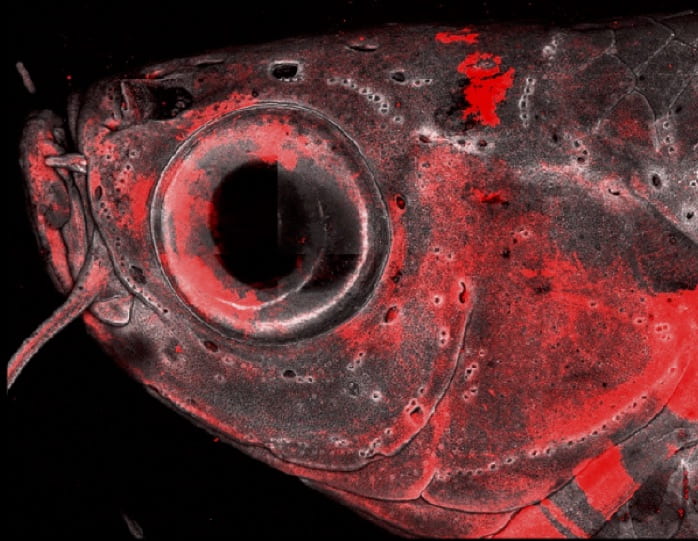
Confocal image of the adult zebrafish head shows neural crest lineage cells in red. (Image courtesy of Peter Fabian)Cranial neural crest cells generate a wide array of mesenchymal cell types in the vertebrate head: chondrocytes, osteocytes, ligament/tendon cells, pericytes, adipocytes, and many more. How are these cell fates chosen in precise spatial locations to build a coherent musculoskeletal system? How do the embryonic progenitors that build the skeleton differ from the adult progenitors that maintain and repair it? By creating a complete catalog of CNC-derived cells across the lifetime of zebrafish, and identifying key regulators of cell fate decisions, we will generate a framework to address these questions. Our approach is to validate hypotheses generated from single-cell genomics approaches with rigorous embryology and genetics in zebrafish.
Modularity and evolutionary modification of craniofacial enhancers
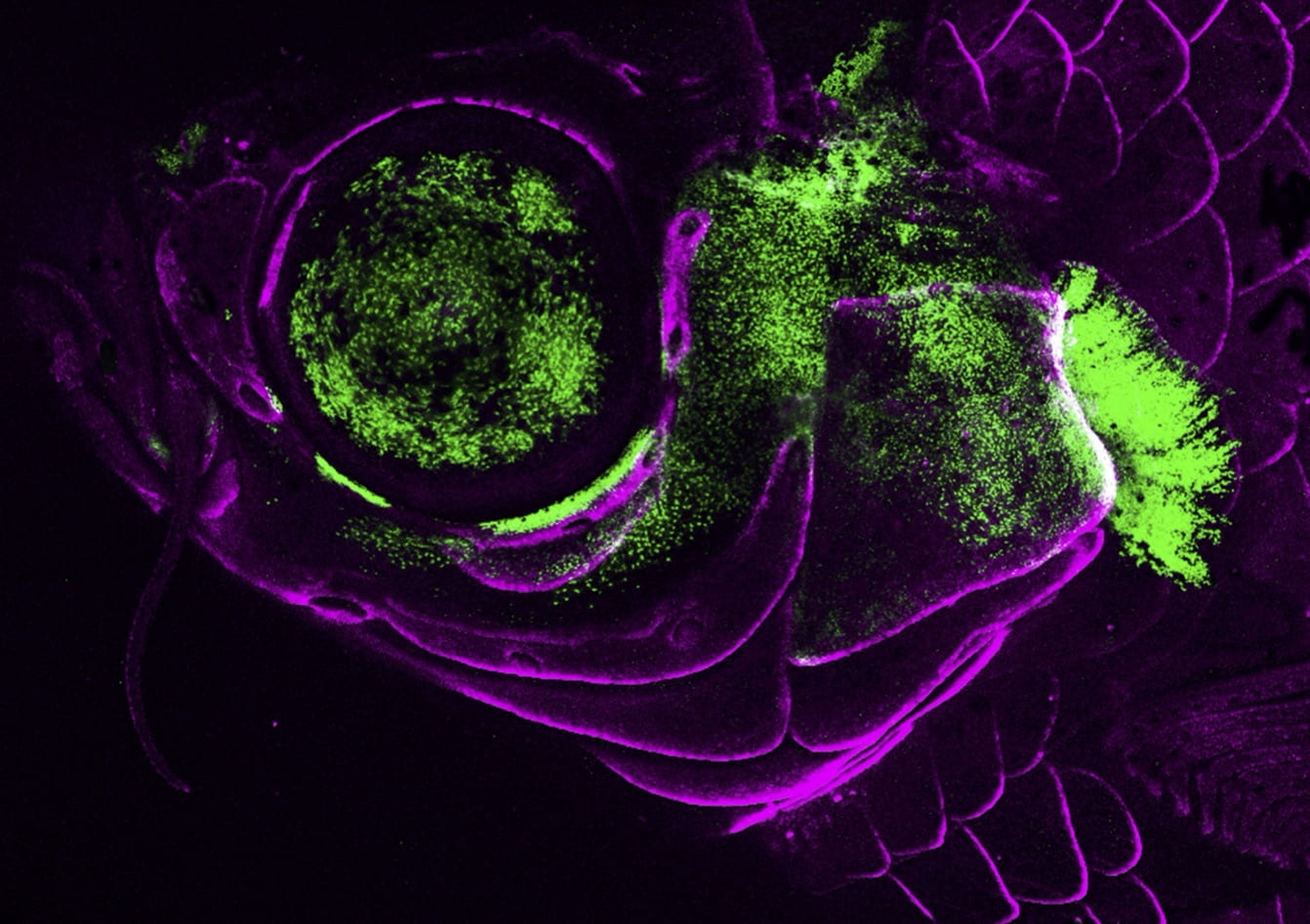
Confocal image of the juvenile zebrafish head shows an ultra-conversed enhancer for the pou3f3b gene driving GFP expression in the gill cover. Bone-forming cells are shown in magenta. (Image courtesy of Lindsey Barske)How is the same cell type specified in different parts of the embryo? How do the functions of related cell types shift during vertebrate evolution? How do new cell types arise? And how does the regeneration of a cell type compare to how that cell type is specified in the embryo? We are tackling these questions by investigating the regulatory elements that promote cell fates in distinct spatial locations, how these regulatory elements are acquired and/or modified during evolution, and how differentiation programs are re-deployed during adult regeneration. Zebrafish has enormous potential for the study of the regulatory non-coding genome. By combining single-cell genomics with transgenic and mutational analysis of putative enhancers in zebrafish, we aim to generate a regulatory logic for the development, evolution, and regeneration of the vertebrate head.
Modeling of human craniofacial birth defects in zebrafish
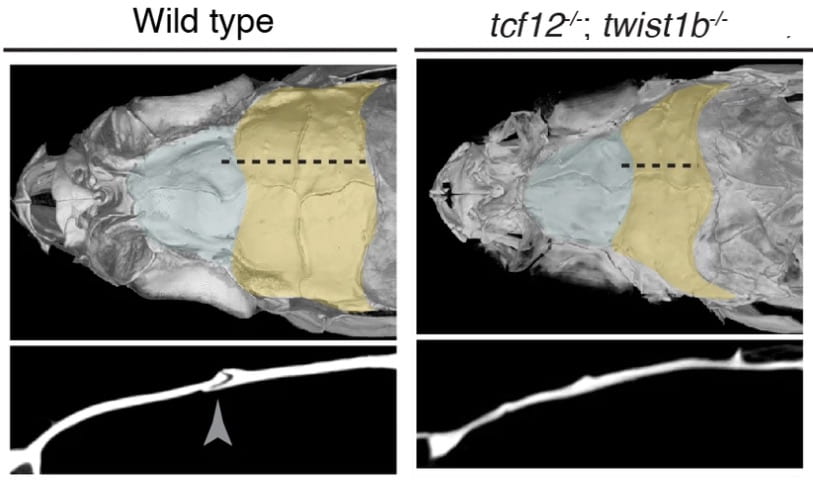
micro-CT images show loss of the suture (see yellow area of skull and arrowhead below) in a zebrafish model of craniosynostosis in Saethre-Chotzen Syndrome. (Image courtesy of Camilla Teng)We are taking advantage of the strong conservation of the cellular and genetic programs between zebrafish and humans to create powerful zebrafish models of human craniofacial birth defects and disease. For example, we have generated zebrafish models for craniosynostosis (Saethre-Chotzen Syndrome), prognathism (Alagille Syndrome), and early-onset arthritis of the jaw (Camptodactyly-arthopathy-coxa vara pericarditis Syndrome). These zebrafish models are revealing how early embryonic misregulation of stem cells can have far-reaching consequences on later maintenance of the head skeleton.
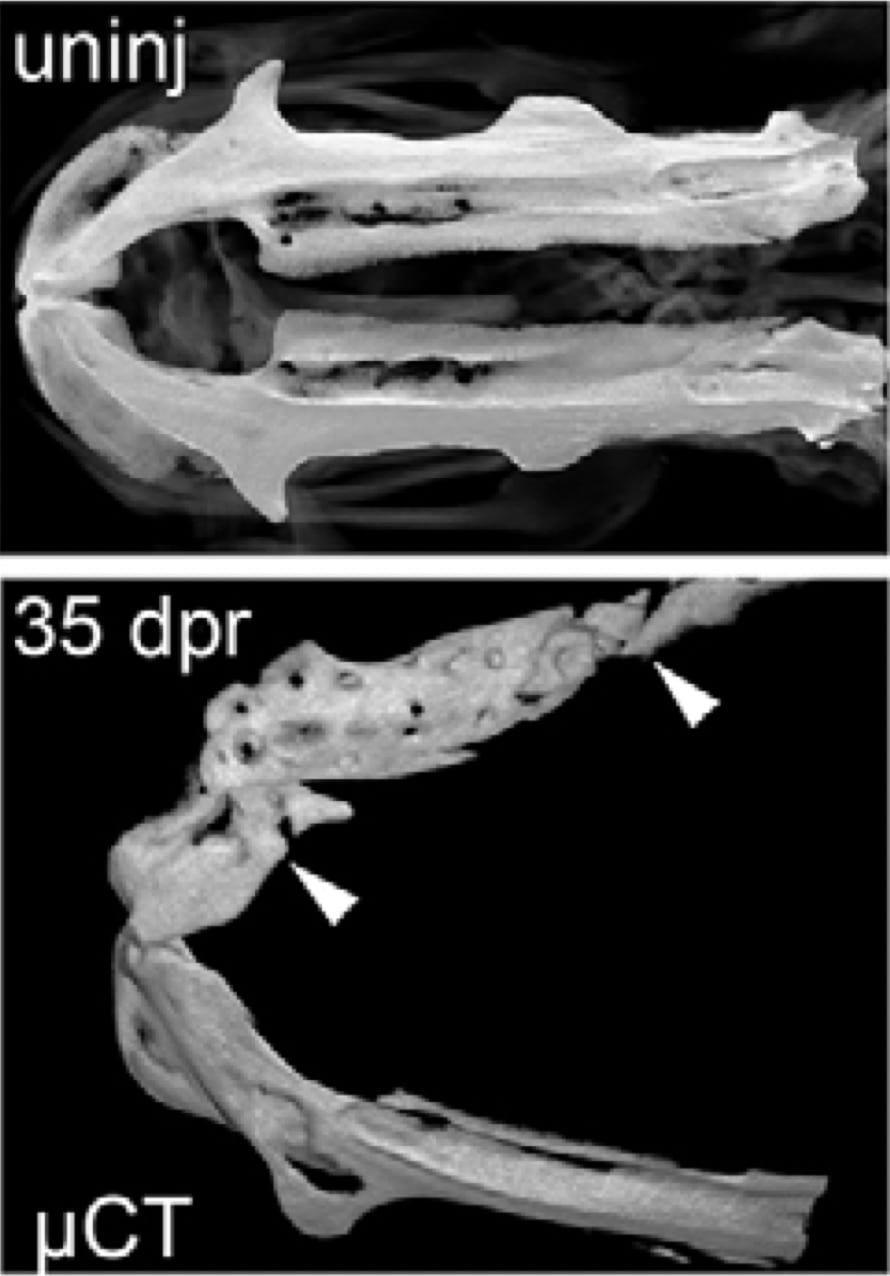
micro-CT images of the lower jawbone in an un-injured zebrafish (above) and 35 days after resection (below). Note the large-scale regeneration of bone in the area between the arrowheads. (Image courtesy of Sandeep Paul)How do adult stem cells detect damaged tissue and replace just the right amount of cells? We have found that zebrafish can robustly regenerate bone, cartilage, and ligament cells in the adult head via two likely origins: pre-existing progenitors and de-differentiating mature cells. We are currently exploring the regulatory logic underlying the striking regenerative capacity of the zebrafish head. Are developmental enhancers maintained in a poised state in adult progenitors, thus ensuring rapid differentiation in response to injury? Do mature cells re-active developmental enhancers upon de-differentiation? Are there regeneration-specific enhancers that respond to general injury cues such as inflammation? And does more robust maintenance of poised lineage enhancers explain the greater regenerative capacity of zebrafish versus mammals? Using single-cell genomics and genetic perturbation approaches, we are contrasting enhancer utilization in regeneration versus development. By better understanding how cell replacement in adults differs from cell generation in embryos, we will be able to develop more targeted therapies for skeletal defects in patients.